
Research Article
Imported Fire Ant (Hymenoptera: Formicidae) Mound Distribution Relationships in Connection to the Heterogeneity of Ten Christmas Tree Farm Landscapes in Mississippi [pdf]
Self, S. R.* and T. E. Nebeker.
Mississippi State University, Department of Entomology and Plant Pathology, Mississippi State, MS 39762
* Corresponding Author: Dept. of Entomology and Plant Pathology, Mississippi State University, P.O. Box 9775, Mississippi State, MS 39762 srd3@entomology.msstate.edu
Received: 8-VII-2010 Accepted: 6-I-2011
Abstract: Ten Christmas tree farms were selected as research sites across Mississippi. At each site a 10-percent inventory was established. Plots were chosen randomly, and all imported fire ant (IFA) (Hymenoptera: Formicidae, Solenopsis spp.) mound positions inside the plot boundaries were captured as global positioning system (GPS) points. Mound points, along with their corresponding volumes (cm3), were recorded and uploaded into ESRI® ArcMap for analysis. Thirteen variables were selected to represent the heterogeneity of these landscapes. Among those variables, the species, height, and diameter of every Christmas tree located inside a plot were recorded, along with average grass height, distance to nearest mound, and other ant species collected within the plot. These data were then analyzed by using analysis of variance and linear regression statistics to determine which variables were significantly related to mound density, mound volume, or both. The statistically significant variables were then subjected to ordinary kriging, a geo-statistical spatial interpolation method. Ordinary kriging was used to illustrate the influence of significant variables in predicting IFA mound distribution. Out of the 13 variables collected and analyzed on each farm, grass height was the only variable significantly related to mound density and volume suitable for use in kriging analysis. All kriging results therefore only included average grass height and average mound volume. For the majority of sites, all larger mounds were clustered in taller grass, while the smaller mounds were scattered across larger areas of shorter grass.
Keywords: Imported fire ants, Solenopsis invicta, Solenopsis richteri, Christmas tree farms, mound distribution
Introduction
Imported fire ants (IFA) (Hymenoptera: Formicidae, Solenopsis spp.) have been the cause of much distress in both rural and urban settings for the past 75 years (Lofgren 1986). Presently, there are two recognized species of IFA in the United States, the black imported fire ant (BIFA), Solenopsis richteri Forel, and the red imported fire ant (RIFA), Solenopsis invicta Buren. Both BIFA and RIFA were accidentally introduced into the port of Mobile, Alabama in the early 1900s, possibly in discarded soil ballast from cargo ships (Vinson 1997). In addition, a hybrid of BIFA and RIFA is found in areas of Alabama and Mississippi where the color phases interface (Vander Meer et al. 1985).
It is estimated that RIFA presently occupy over 10,440,000 ha, which, based on their introduction to Alabama in the mid-1930s, indicates a westward migration of approximately 19.8 km/yr (Vinson 1997). Long-distance distribution of whole colonies has been facilitated by inadvertent spread through nursery stock, soil, sod, on road construction equipment, through pipelines, electrical and telecommunication lines, and in landfill movement. Natural mating flights can also land reproductive females in open pickup trucks, railroad cars, and trailers that might transport the colonies for hundreds of miles (Vinson 1997, Taber 2000).
The first sign of fire ant colonies is often their nest mounds, where their brood is reared. Mounds may reach 28–38 cm in height (Vinson 1997) depending on soil type. The actual mound is the above-ground portion of the nest where the brood is reared. The workers can distinguish changes in temperature and humidity in this area of the nest, and move the brood based upon the ideal environment for their development (Taber 2000). A single mound consists of many brood-rearing chambers and foraging tunnels radiating from the center of the mound (Wilson et al. 1971). The foraging tunnel system is extensive, and allows workers to forage during parts of the day when soil surface temperatures would be lethal (Porter & Tschinkel 1987).
Habitats usually preferred by RIFA include state parks, commercial orchards, and especially open pasture land (Taber 2000). Diffie and Bass (1994) found that colony densities in Georgia pastures averaged about 27 mounds per 0.405 ha. The same amount of recently disturbed land might harbor more than a thousand colonies (Lofgren et al. 1975). Mounds are also found in other open or disturbed areas such as clear-cuts, re-planted forests, margins of seasonal ponds, along vehicle tracks, and in and around human constructs. According to Buren (1972) and Stiles and Jones (1998), RIFA are able to dominate in these types of altered habitats. This would seem to be proven by the many cases of their unique ability to withstand disturbances and even flourish under adverse conditions. Russellet al. (2001) reported that highway roadbeds were frequently infestated with RIFA. RIFA prefer early successional communities (Tschinkel 1986) and can efficiently out-compete other ant species in ecologically disturbed habitats (Urbani & Kannowski 1974, Tschinkel 1988). RIFA reproduce early and at high rates, they have developed an efficient method of mass dispersal, and they specialize at colonizing and dominating disturbed habitats (Taber 2000). While several studies have looked at the distribution of IFA mounds in pastureland and farm settings (Vogt et al. 2004, Vogt et al. 2009) Christmas tree plantations have been relatively overlooked, despite the fact they provide IFA with prime early successional habitat. They undergo frequent disturbance due to mowing regimens and site preparation, and many have open, sunny areas where trees have been harvested.
Christmas trees come from thousands of evergreen plantations across the United States. Over 350,000 ha of land have been planted in Christmas trees, providing fire ants with ample acreage for infestation (Wolford 2011). Midsouth farmers supply a very modest market compared to northern locales, but their economic impact is still important. Rather than selling mass quantities of trees to ship to commercial businesses, most plantations in the South are “choose and cut” farms, where the buyer is allowed to walk through the growing trees, select a tree, and cut it. Mississippi currently has fewer than 100 farms, which sell an average of 1,000 trees/farm/year. Alabama and Louisiana have slightly more choose and cut farms, though their production levels, cultural practices, and IFA infestation rates are very similar to those found in Mississippi (Drapala 2008, Wolford 2011).
Christmas trees should be viewed as a sustainable resource, not unlike cotton or soybean crops in the southeastern United States. Approximately 73 million new Christmas trees are planted annually, with over two seedlings re-planted for every tree harvested. In Mississippi the trees are generally grown on a four-year rotation, as compared to the seven to ten year rotations needed in colder, northern climates (Wright 2002). Common species grown in the South include: Virginia pine (Pinus virginiana), Leyland cypress (X Cupressocyparis leylandii), Carolina Sapphire (Cupressus arizonica), and eastern red cedar (Juniperus virginiana). Basic cultural practices utilized by Christmas tree growers include various methods of site preparation, planting, grass control, shearing (trimming branches into “Christmas-tree-shape”), and pest control (Wright 2002). The intensity of these practices varies greatly from farmer to farmer, and can consist of simply planting the trees and letting them grow, to a rigid schedule of mowing, shearing, and herbicide/pesticide applications.
Christmas tree plantations provide IFA with the early successional habitats they prefer for infestation and colonization. Therefore, infestation levels in these areas can be very high, presenting growers with heavy worker and customer liability, and equipment damage. Although not as common as the liability and equipment problems, IFA also cause seedling damage by building large mounds at the base of young trees, eventually covering a large portion of the trunk, causing stunted growth and deformed shape (Wright 2000). The objective of the study presented here was to identify variables within Christmas tree farms that represented the heterogeneity of those landscapes, and then use those variables in conjunction with geo-referenced IFA mounds to interpolate mound densities and distributions by using kriging techniques. Variables determined to be statistically significant factors in mound density and volume were subjected to ordinary kriging to predict mound distribution throughout farm landscapes. This information could potentially help farmers avoid or decrease high mound densities, which in turn would reduce possible equipment damage, worker and customer liability, and seedling damage. Only those variables found to be statistically significant in relation to mound distribution were discussed in the portion of the study presented here.
Materials and Methods
Ten Christmas tree farms, located across the state of Mississippi, were selected as research sites (Table 1). The locations were spread across Mississippi to account for any differences that might exist between physiographic regions of the state. All sites were subjected to varying levels of rainfall and other climatic differences throughout the study period. Using sites located in different physiographic regions eliminated possible bias that might have been present if all sites had been located in one particular area. All landowners selected belonged to the Southern Christmas Tree Association (SCTA), and participated in a preliminary written survey used to identify various cultural practices used on each farm. These variables were not used in the kriging analyses, due to an absence of x, y coordinates for each. Kriging analyses require an x, y component for each variable, therefore only plot-level data, rather than data—such as mowing schedules—collected for the entire farm, could be used to analyze and predict mound distribution geospatially.
Table 1. Site number and corresponding location and physiographic region.
Site Number |
County |
Physiographic Region |
1 |
Alcorn |
Upper Coastal Plain |
2 |
Coahoma |
Delta |
3 |
Panola |
Brown Loam/Thick Loess |
4 |
Lee |
Blacklands |
5 |
Lowndes |
Blacklands |
6 |
Attala |
Upper Coastal Plain |
7 |
Rankin |
Thin Loess |
8 |
Jones |
Lower Coastal Plain |
9 |
Pike |
Lower Coastal Plain |
10 |
Pearl River |
Coastal Flatwoods |
All farms consisted of approximately 1.6 ha or less and grew at least two different species of Christmas trees, with the exception of Site 7 in Rankin County, which specialized in Leyland cypress. All measurements were collected in the summers of 2003, 2004, and 2005, between mid-May and mid-August, when IFA mound building is generally at its peak. The farms varied widely in cultural practices and in planting mosaics. Due to this variability, one 0.04-ha plot was established per 0.40 ha present, providing a 10% inventory of the entire plantation. Plots were chosen entirely at random. Plot centers and all IFA mounds present in the plot boundaries were geo-referenced by using an Allegro CETM Global Positioning System (GPS) expansion pod (Juniper Systems Inc., Logan, UT). A perimeter around the plantation was also geo-referenced with the Allegro unit.
Within each plot tree species, diameter, and height were recorded with the aid of an Allegro data logger. Diameter was measured to the nearest 0.1 mm by using a standard digital caliper for seedlings, or to the nearest 2.54 mm by using a standard forestry caliper for trees equal to or greater than 1.5 m in height. Diameter at breast height (DBH) was measured on trees 1.5 m and above, otherwise, ground-line diameter (GLD) was measured at the base of the tree. Total height was measured to the nearest 0.03 m by using a Crain® telescoping leveling rod. Spacing between trees and between rows was measured and recorded by using a standard loggers tape. Grass height was measured at plot center on every plot. All species of grasses were considered similar for this portion of the study, with height, not individual species, being the statistically important variable. Width, length, and height of each mound was measured and recorded to the nearest 0.1 mm and used to calculate mound volume based upon the formula given in Macom and Porter (1996). If a mound was not raised above ground level, a standard height of 0.61 cm was recorded, as recommended by Macom and Porter (1996). Since IFA mounds can be connected to one another, distance to the nearest mound was measured on each plot for each IFA mound within the plot. Nearest mounds did not have to be located within the plot to be counted, and were measured to the nearest 2.54 mm.
A digital orthophoto quarter quadrangal (DOQQ) of each study site, based on aerial photography taken during the winters of 1995 and 1996, was obtained from the Mississippi Automated Resource Information System (MARIS) server (http://www.maris.state.ms.us/). All DOQQs have a ground resolution of one meter, and have Mississippi transverse mercator (MSTM) coordinates that correspond to USGS 7.5 minute, 1:24,000 topographic maps. All DOQQs were used to create maps of the farms and to display the mounds that were geo-referenced.
ESRI ArcMap® (ArcGIS version 10, ESRI, Redlands, CA) was used to perform the geostatistical analyses. These methods were used to determine what distributional relationships existed between the mounds themselves, and between the mounds and any statistically significant variable that was measured in the landscape. Data were analyzed for significance by using Proc ANOVA and Proc REG (SAS Institute, version 8.0). Kriging was determined to be the most accurate method for geostatistical interpolation, because, as a predictor, it does not require the data to be normally distributed as do inverse distance-weighted (IDW) methods. Ordinary kriging was used rather than universal kriging, block kriging, or cokriging. Ordinary kriging assumes the absence of a “drift,” or structure representing a trend, and focuses instead on spatially correlated components. The other methods of kriging all incorporate a second-order polynomial in the kriging process to represent possible drift or trend (Isaaks & Srivastava 1989).
Results and Discussion
General Results. Two farms from northern Mississippi (Site #1 and Site #4) were excluded from this part of the study due to a lack of IFA presence. This absence could be due to a naturally unsuitable habitat, high levels of control, or both. Site #1 was located on the second highest elevation in Mississippi (241 m), and grew primarily eastern white pine (Pinus strobus), which is atypical for the South. Site #4 was completely surrounded by mature mixed pine/hardwood forest, which is not typically preferred habitat for IFA. Additionally, this farm was also subjected to frequent insecticide applications, unlike any other farm.
Out of the 13 variables collected and analyzed on each farm, grass height was the only variable significantly related to mound density and volume that could be used in the kriging analysis (Table 2). Kriging analyses require an x, y component for each variable, therefore only plot-level data, rather than data collected for the entire farm (such as fertilization schedule) could be used to analyze and predict mound distribution geospatially. Average mound volumes were calculated on a plot-level basis, and are reported per site in Table 3. These numbers were used in predicting mound distribution as a function of average grass height by using ordinary kriging techniques.
Table 2. Regression results for plot level effects on mound volume. |
|||
Effect |
Estimate |
F |
Pr > F |
Number of Trees |
-0.0322 |
0.2 |
0.6572 |
Grass Height |
0.0332 |
3.87 |
0.0503 |
Table 3. Average IFA mound volume (cm3) for each site. |
||
Site # |
Mean |
Standard Deviation |
1 |
0 |
0 |
2 |
3378.43 |
8264.06 |
3 |
5437.43 |
7284.97 |
4 |
0 |
0 |
5 |
9263.48 |
127777.6 |
6 |
3243.88 |
4402.9 |
7 |
2901.45 |
4576.62 |
8 |
1894.91 |
2080.95 |
9 |
6291.95 |
5904.62 |
10 |
1866.46 |
2765.2 |
Kriging Analysis Results. Figure 1A and 1B serve as general examples of kriging analysis results produced in ESRI ArcMap®. The predicted distribution of IFA mounds based on grass height for Site #10 is shown in Figure 1A. The red dots are actual ant mounds in the field, and the strips of color represent various grass heights. The width of the strip represents the distribution pattern of the mounds within that particular range. Higher mound numbers were predicted for shorter grass zones. Interestingly, these were also for the smallest mound volumes. Figure 1B represents the distribution pattern of the mounds based upon their average mound volumes. The largest mounds were located in the areas of the tallest grass. Sites #3, #6, #7, and #8 (Panola, Attala, Rankin, and Jones Counties, respectively) all illustrated similar patterns for their corresponding locations. Theoretically, this relationship is very reasonable. Showler and Reagan (1987) stated that IFA prefer taller grasses for mound construction because the vegetation provides support and structure as the mounds increase in size. Although not statistically significant in this study, it is probable that the shorter grass was a result of mowing schedule, which would consequently increase mound disturbance. This disturbance might have kept the mounds below a certain threshold size, increasing the significance of the less disturbed, taller grasses in areas not mowed, such as around stumps and under tree limbs. Mowing schedule was one of the 13 variables analyzed, but, probably due to the wide variation between farms, it was found to not be significantly related to average mound volume or distribution in the study.
The same result was not found for Site #2 in Coahoma County. More mounds were located, once again, in the shorter grass ranges, but the average mound volume did not follow the pattern expressed in the above farms. This farm was located in the delta region of Mississippi, and sustained a large population of RIFA. Unfortunately, on the day of the first survey, half of the farm had recently been bush-hogged, while the remaining half had not. Consequently, many of the mounds measured for volume were much flatter than they would otherwise have been. Half of the mounds were recorded as having large volumes, while the other half was recorded as having very reduced volumes. This discrepancy was not representative of the actual situation on the farm. In better circumstances, the mound volume distribution would probably have mimicked those of the other farms previously mentioned.
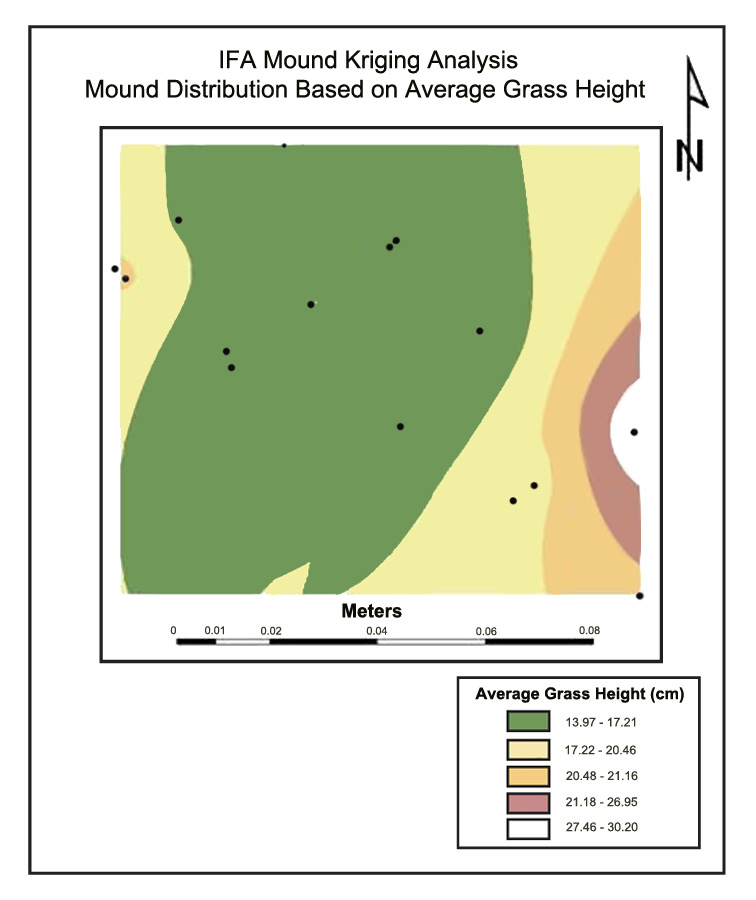
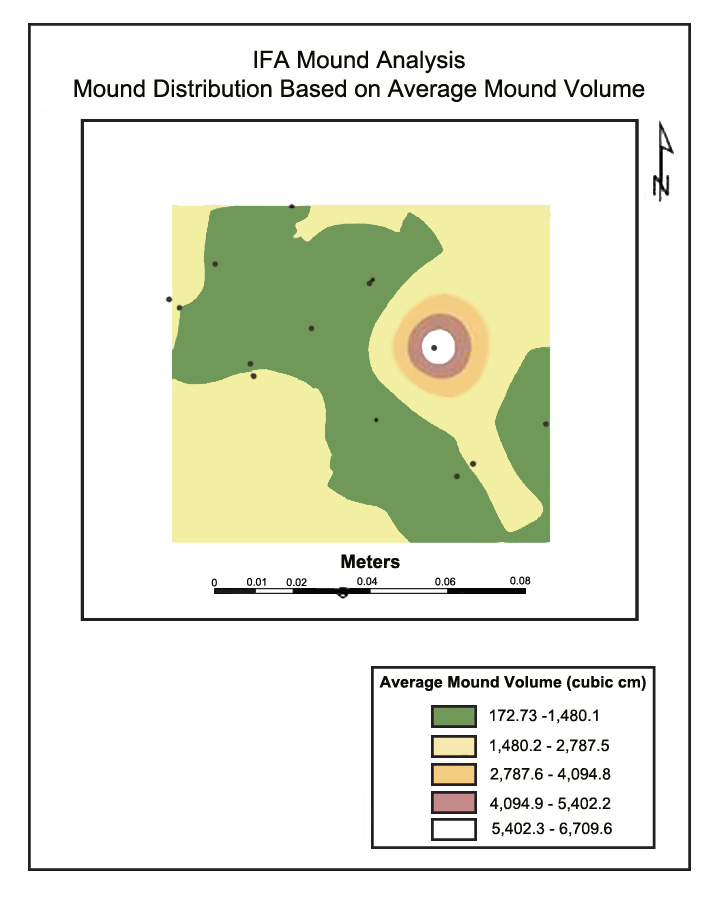
Figure 1. (A) Kriging results for grass height and (B) mound volume on Site #10, Pearl River County, MS, 2005.
Site #9 in Pike County displayed different results as well. In the average grass height analysis, there was a much larger section showing taller grass, and therefore a greater abundance in IFA mounds. Interestingly, these mounds, according to the mound volume analysis, correspond, for the most part, with “medium” sized mounds. This farm had an abundance of average mounds, regardless of the grass height. A possible explanation for this phenomenon might have been rainfall events occurring several days prior to data collection. Excessive rains may have temporarily decreased mound heights until the colony was able to rebuild mounds to original heights. August 2005 was exceptionally rainy, especially for southern counties such as Pike County, which experienced the effects of Hurricane Katrina.
The analysis for Site #5 in Lowndes County indicated that the entire farm had one average grass height. The largest volumes were predicted on the east side of the property, where tree harvesting had not taken place in several years, and was therefore rarely disturbed. This part of the farm was also located next to a sunflower field, unlike any of the other research sites. Lofgren et al. (1975) reported RIFA foraging in sunflowers, so it is possible that the presence of a preferred food source could have skewed the data collected. Herbaceous vegetation was surveyed on all research sites, but was found to have no statistical significance in relation to mound volume or distribution, eliminating other plants located on the farm as a possible explanation. Additionally, Reinert (2010), and Reinert and McCoy (2010) reported RIFA feeding preferences for different turfgrass cultivars. They found that Bermudagrass (Cynodon sp.) was preferred over all other grass species tested. Grass species were recorded for all ten research sites, and were found to have no statistical significance in relation to mound volume or distribution. However, all sites had over 90% Bermudagrass present. The study did not differentiate between different cultivars.
The study identified 13 variables, including tree height, diameter and species, ant species present, herbaceous vegetation present, distance to nearest mound, and various cultural practices including fertilization schedule and mowing schedule, that could be used to describe landscape heterogeneity. Out of these 13 variables, only fertilization schedule and average grass height were found to be statistically significant in predicting mound density and size, (p = 0.05 and 0.1). These two factors might be related due to fertilization schedules impacting grass heights observed on site. Additionally, the study presented here was limited to a plot level analysis, because x, y coordinates were needed to complete the prediction formula in the geospatial prediction analysis. Therefore, the only variable available for ordinary kriging was average grass height because fertilization schedule information was obtained for the entire farm, rather than for individual plots.
Kriging methods predicted, for five of the eight sites analyzed, that mature IFA colonies prefer taller grasses in which to build their mounds. This might indicate that colonies prefer sites that are less frequently disturbed. Whether or not the smaller mounds were new colonies, foraging tunnel outlets, or simply frequently disturbed mounds is unknown. Distance to nearest mounds was analyzed, since it is likely that at least some of the mounds were connected. However, this variable was found to have no statistical significance in this particular study. Whatever the situation, ordinary kriging analyses consistently interpolated the presence of smaller mounds in shorter grasses where they were more frequently disturbed. The only exceptions to this pattern were three farms, sites #2, #5 and #9 (Coahoma, Lowndes, and Pike Counties, respectively) that had extenuating circumstances surrounding either the collecting dates, poorly timed bush-hogging, adjacent preferred food sources (sunflower fields), or inclement weather that could possibly have skewed the data and therefore the resulting analyses. It is plausible that, had the data been collected under better circumstances, these sites would have also shown similar interpolation patterns.
Mound distribution was also predicted based on mound volumes present. This was done for comparison with the previous analysis, and also for additional description of landscape heterogeneity. In most cases, the majority of mounds were small to medium in volume, with only a few very large mounds present. This circumstance was expected, and has been noted by other studies including Byron and Hays (1986) and Diffie and Bass (1994), who found that mature IFA colonies typically send out reproductive pairs during nuptial flights that can start multiple, smaller colonies nearby.
The ability to predict distribution patterns of IFA mounds, as a function of common cultural practices and landscape heterogeneity, could have the potential to help farmers avoid or decrease high mound densities, which in turn would reduce possible equipment damage, worker and customer liability, and seedling damage. This study was limited to a plot-level analysis, due to the qualitative nature of the written survey used to identify landscape level variables. However, future work could easily be pursued by applying the identified variables as controlled treatments in georeferenced locations.
Acknowledgements
Approved for publication as Journal Article No. J-11825 of the Mississippi Agricultural and Forestry Experiment Station, Mississippi State University. Thanks for field assistance to Mr. Andrew Self, Mr. Michael Guttery, Mr. Jeremy Norris, and Mr. Keith Beatty. Thanks for GIS assistance to Dr. Cook, Dept. of Geosciences, Mississippi State University.
References
Buren, W. F. 1972. Revisionary studies on the taxonomy of the imported fire ants. J. Georgia Entomol. Soc. 7: 1-26.
Byron, D. W. and S. B. Hays. 1986. Occurrence and significance of multiple mound utilization by colonies of the red imported fire ant (Hymenoptera: Formicidae). J. Econ. Entomol. 79: 637-640.
Diffie, S., and M. H. Bass. 1994. Densities of monogynous red imported fire ant (Hymenoptera: Formicidae) colonies in Georgia pastures. J. Entomol. Sci. 29: 367-369.
Drapala, P. 2008. Christmas tree crop shapes up to market. MSU Extension Fact Sheet 11-20-2008. On-line posting (http://msucares.com/news/print/fwnews/fw08/081120trees.html) accessed 15 February, 2011.
Isaaks, E. H., and R. M. Srivastava. 1989. Introduction to Applied Geostatistics. Oxford University Press, New York, NY. 553 pp.
Lofgren, C. S., W. A. Banks, and B. M. Glancey. 1975. Biology and control of imported fire ants. Ann. Rev. Entomol. 20: 1-30.
Lofgren, C. S. 1986. History of the imported fire ants in the United States. pp 36-47 In: Lofgren and Vander Meer (eds.) Fire Ants and Leaf-Cutting Ants Biology and Management. Westview Press, Boulder, CO.
Macom, T. E., and S. D. Porter. 1996. Food and energy requirements of laboratory fire ant colonies (Hymenoptera: Formicidae). Environ. Entomol. 24: 387-391.
Porter, S. D., and W. R. Tschinkel. 1987. Foraging in Solenopsis invicta (Hymenoptera: Formicidae): effects of weather and season. Environ. Entomol. 16:1987.
Rhoades, R. B. 1977. Medical Aspects of the Imported Fire Ant. Gainsville: University Press of Florida. 75pp.
Reinert, J. A. 2010. Red imported fire ants forage on turfgrass. USGA Turfgrass and Environmental Research Online 9(20): 1-9.
Reinert, J. A. and J. E. McCoy. 2010. Foraging by red imported fire ants, Solenopsis invicta (Hymenoptera: Formicidae) on turfgrass. Florida Entomol. 93: 248-253.
Russell, S. A., H. G. Thorvilson, and S. A. Phillips Jr. 2001. Red imported fire ant (Hymenoptera: Formicidae) populations in Texas highway rights-of-way and adjacent pastures. Environ. Entomol. 30: 267-273.
SAS Institute, Inc., 1999. SAS Verion 8.0. Cary, NC.
Showler, A. T. and T. E. Reagan. 1987. Ecological interactions of the red imported fire ant in the southeastern United States. J. Entomol. Sci. Suppl. 1: 52-64.
Stiles, J. H., and R. H. Jones. 1998. Distribution of the red imported fire ant, Solenopsis invicta, in road and powerline habitats. Landscape Ecology 13: 335-346.
Taber, S. W. 2000. Fire Ants. Texas A&M University Agriculture Series, Number Three. Texas A&M University Press, College Station, TX.
Tschinkel, W. R. 1986. The ecological nature of the fire ant: some aspects of colony function and some unanswered questions. pp. 72-87 in Lofgren and Vander Meer (eds.) Fire Ants and Leaf-Cutting Ants Biology and Management. Westview Press, Boulder, CO.
Tschinkel, W. R. 1988. Distribution of the fire ants Solenopsis invicta and S. geminata (Hymenoptera: Formicidae) in northern Florida in relation to habitat and disturbance. Ann. Entomol. Soc. Am. 81: 76-81.
Urbani, B. C., and P. B. Kannowski. 1974. Patterns in the red imported fire ant settlement of a Louisiana pasture: some demographic parameters, interspecific competition and food sharing. Environ. Entomol. 3: 755-760.
Vander Meer, R. K., C. S. Lofgren, and F. M. Alvarez. 1985. Biochemical evidence for hybridization in fire ants. Florida Entomol. 68: 501-506.
Vinson, S. B. 1997. Invasion of the red imported fire ant (Hymenoptera: Formicidae): spread, biology, and impact. Amer. Entomol. 43: 23-39.
Vogt, J. T., D. A. Streett, and D. Boykin. 2004. Seasonal characteristics of black imported fire ant (Hymenoptera: Formicidae) mounds in northern Mississipi pastures. Sociobiology 43: 513-522.
Vogt, J. T., M. L. Allen, B. Wallet, D. Boykin and W. A. Smith. 2009. Distribution patterns of imported fire ants (Hymenoptera: Formicidae) on a sheep and goat farm in Oklahoma. Environ. Entomol. 38: 551-560.
Wilson, N. L., J. H. Dillier, and G. P. Markin. 1971. Foraging territories of imported fire ants. Ann. Entomol. Soc. Am. 64: 660-665.
Wolford, R. 2011. Christmas Tree Facts, Christmas Trees & More, University of Illinois, On-line posting (http://urbanext.illinois.edu/trees/facts.cfm) accessed 15 February, 2011.
Wright, C. W. (ed.). 2002. Christmas Trees from Seed to Sale. Tree Publishers Inc, Lecompton, KS. 43 pp.